Сознание
– действительно ли «за пределами мозга»?
И.В. Лимарь
(Получена 13 августа 2009; изменена 20 августа 2009;
опубликована 15 октября 2009)
В статье рассматриваются некоторые спорные вопросы психофизиологии,
поднимавшиеся различными исследователями и возможность их актуализации с учетом
новейших достижений физики квантовой информации
Название книги известного американского психолога чешского происхождения
Станислава Грофа «За пределами мозга» [13], как нельзя более точно отражает
суть проблем, затрагиваемых в настоящей публикации. Однако, увы – история вопроса
гораздо более обширна и не ограничивается теорией основателя трансперсональной
психологии С. Грофа.
Итак, по порядку. В свое
время крупнейший современный нейрофизиолог, лауреат
Нобелевской премии по медицине Дж. Экклз развил мысль о том, что на основе
анализа деятельности мозга невозможно выяснить происхождение психических
явлений, и этот факт легко может быть истолкован в том смысле, что психика
вообще не является функцией мозга [58]. Со своим коллегой, основоположником
современной нейрохирургии Уайлдером Пенфилдом, выполнившим свыше 10 000
операций на мозге, Экклз написал книгу «Тайна человека». В ней авторы прямо
заявляют, что «нет никаких сомнений в том, что человеком управляет НЕЧТО, находящееся
за пределами его тела». Экклз пишет, что «работа сознания не может быть
объяснена функционированием мозга. Сознание существует независимо от него
извне». Уилдер Пенфилд в результате многолетнего изучения деятельности мозга
тоже пришел к выводу, что «энергия разума отличается от энергии мозговых
нейронных импульсов» [57]. Академик Академии медицинских наук РФ, директор
Научно исследовательского института Мозга (РАМН РФ), нейрофизиолог с мировым
именем , профессор, д.м.н. Наталья Петровна Бехтерева: «Гипотезу о том, что
мозг человека лишь воспринимает мысли откуда-то извне, я впервые услышала из
уст нобелевского лауреата, профессора Джона Экклза. Конечно, тогда это
показалось мне абсурдным. Но потом исследования, проводимые в нашем
Санкт-Петербургском НИИ мозга, подтвердили: мы не можем объяснить механику
творческого процесса. Мозг может генерировать лишь самые простые мысли типа,
как перевернуть страницы читаемой книги или помешать сахар в стакане. А
творческий процесс — это проявление совершенно нового качества». Профессор С.
Гроф так говорит об этом: «представьте, что у вас сломался телевизор, и вы
вызвали телемастера, который, покрутив разные ручки, его настроил. Вам же не
приходит в голову, что все эти станции сидят в этом ящике».
Голографическая картина мира
В последние годы жизни Дэвид Бом - учёный, внесший вклад в квантовую
физику, философию и нейропсихологию, много времени уделял работе над
голографической моделью Вселенной. Бом считал, что все индивидуумы
взаимосвязаны не в результате непосредственного влияния, которое они могут
оказывать друг на друга, а в силу того факта, что все они подвержены влиянию
общих фундаментальных законов. По Бому все, включая сознание и материю, активно
влияет на целое, а посредством целого и на все составляющие [7].
В своей теории «голодвижения» (holomovement) Бом предполагал, что каждый
пространственно-временной участок мира содержит в себе весь порядок вселенной.
Это включает в себя, как прошлое, так настоящее и будущее. Подобно голограмме,
где каждый сегмент содержит информацию о целом запечатленном объекте, каждый
участок воспринимаемого нами мира содержит в себе полную информацию о структуре
вселенной, или целого мира. В этой холистской концепции никто и ничто в нашей
жизни не остров. Все, включая мысли и поступки, произрастает из единой основы,
приводя к тому, что любое изменение в одной части мира, немедленно
сопровождается или отражается в соответствующих изменениях во всех остальных
частях. Эти теории Бома были использованы американским нейропсихологом Карлом
Прибрамом, рассматривавшим мозг как голографическую структуру [38].
И, наконец, следует привести высказывание
Карла Густава Юнга – основателя такого направления глубинной психологии как
«аналитическая психология»: «Мы должны полностью отказаться от идеи, что
психика каким-то образом связана с мозгом. Мы оказываемся гораздо ближе к
первичному фактору, который, не имеет ничего общего с деятельностью мозга» [56].
Здесь не будет обсуждаться теория «коллективного бессознательного» - одна из
основ учения К. Г. Юнга. Это тема для отдельной статьи. А вот о том, что гипотеза
«синхронии» - смыслового совпадения поведения людей без видимых причинных
связей являлась предметом пристального внимания не только самого К. Г. Юнга, но
и тесно с ним сотрудничавшего известного физика, работавшего в области
квантовой механики В. Паули, следует упомянуть.
Каковы
же могут быть физиологические (и физические) механизмы лежащие в основе столь
ошеломительных предположений ?
После
постановки группой французских ученых, под руководством Элайна Аспекта,
эксперимента по проверке неравенств Белла прошло двадцать восемь лет. За это
время усовершенствовалась лабораторная база, и было поставлено много других
впечатляющих экспериментов, положивших начало новому направлению в физике – квантовой
информатике. Одним из основных положений квантовой теории информации является,
как известно феномен квантовой нелокальности или квантовой запутанности: два «объекта»
микромира один раз «побывав в контакте» ( если слова в кавычках вообще
применимы к явлениям микромира ) становятся «связанными» общей волновой
функцией и, впоследствии, будучи разнесенными на сколь угодно большие
расстояния, подвержены взаимным корреляциям, причем, мгновенно, без ограничений
по скорости накладываемых специальной теорией относительности. Правда свои
коррективы в данный феномен вносит явление декогеренции - исчезновение
наблюдаемых квантовомеханических свойств
системы при «взаимодействии» с макроокружением. Но об этой проблеме в статье
будет изложено далее и более детально.
Таким
образом, перед нами очевидный кандидат для объяснения изложенных в начале
статьи гипотез.
Но
физиологические процессы в организме обусловлены не исключительно физической
формой движения материи, а стоящими на более высоких ступенях иерархии ( как мы
это изучали в курсе философии ) химической и биологической. Что ж – обратимся вначале к основным
положениям квантовой химии, а далее – к молекулярной биологии.
Ключевые понятия квантовой химии
Электронные облака - орбитали
Единственный
электрон атома водорода образует вокруг ядра сферическую орбиталь -
шарообразное электронное облако, вроде неплотно намотанного клубка пушистой
шерсти или ватного шарика [11].
Сферическую атомную орбиталь ученые
договорились называть s-орбиталью. Она самая устойчивая и
располагается довольно близко к ядру.

Чем больше энергия электрона в
атоме, тем быстрее он вращается, тем сильнее вытягивается область его
пребывания и наконец превращается в гантелеобразную
p-орбиталь:
 Электронное облако такой формы может занимать
в атоме три положения вдоль осей координат пространства x, y
и z. Это легко объяснимо: ведь все электроны заряжены
Электронное облако такой формы может занимать
в атоме три положения вдоль осей координат пространства x, y
и z. Это легко объяснимо: ведь все электроны заряжены 
 отрицательно, поэтому
электронные облака взаимно отталкиваются и стремятся разместиться как
можно дальше друг от друга.
отрицательно, поэтому
электронные облака взаимно отталкиваются и стремятся разместиться как
можно дальше друг от друга.
Все вместе три
электронных облака, которые называют px-, py-
или pz-орбиталями, образуют симметричную геометрическую
фигуру, в центре которой находится атомное ядро. Она похожа на шестиконечный
помпончик или на тройной бант - кому как нравится.
Итак, p-орбиталей может быть
три. Энергия их, конечно, одинакова, а расположение в пространстве - разное.
Кроме
s- и p-орбиталей, существуют электронные орбитали еще более
сложной формы; их обозначают буквами d и f. Попадающие сюда
электроны приобретают еще больший запас энергии, двигаются по сложным путям, и
в итоге получаются сложные и красивые объемные геометрические фигуры.
Все d-орбитали (а их
может быть уже пять) одинаковы по энергии, но по-разному расположены в
пространстве. Да и по форме, напоминающей перевязанную лентами подушечку,
одинаковы только четыре. А пятая - вроде гантели, продетой в бублик.
При
сближении двух атомов водорода, содержащих электроны с параллельными спинами,
энергия системы постоянно увеличивается и связь не образуется.
При
наличии у пары электронов противоположных спинов электроны двигаются в поле
обоих ядер. Между ядрами появляется область с высокой плотностью электронного
облака – избыточного отрицательного заряда, который стягивает положительно
заряженные ядра.
Связь образуется электронной парой с
антипараллельными спинами. Прочность
связи пропорциональна степени перекрывания электронных облаков.
Направленность связи обусловлена
перекрыванием электронных облаков в области максимальной электронной плотности.
Для
молекул более сложных, принципы расчета остаются неизменными. К образованию
связи приводит взаимодействие пары электронов с противоположными спинами, но с
волновыми функциями одинакового знака, которые суммируются. Результатом этого
является увеличение электронной плотности в области перекрывания электронных
облаков и стягивание ядер.
Наибольшая
плотность электронного облака у 2р-орбитали в направлении оси симметрии. Если
неспаренные электроны атомов фтора находятся на 2рх-орбиталях, связь
осуществляется в направлении оси х .

Образование молекулы F2
В
молекуле фтороводорода НF связь образована 1s-орбиталью атома водорода и 2рх-орбиталью
атома фтора.
Направленность
связи в этой молекуле определяется ориентацией 2рх-орбитали атома фтора.
Перекрывание происходит в направлении оси симметрии х. Любой другой вариант
перекрывания энергетически менее выгоден.

Образование молекулы НF
Более
сложные d- и f-орбитали также характеризуются направлениями максимальной электронной
плотности вдоль осей их симметрии.
Поскольку
оси симметрии валентных 3р-орбиталей атома серы взаимно перпендикулярны, то
следует ожидать, что молекула Н2S должна иметь уголковую структуру с
углом между связями S–Н 90°. Действительно, угол близок к расчетному и равен
92°.

Образование молекулы Н2S
Пара
электронов с антипараллельными спинами, образующая связь, может быть получена
не только по обменному механизму, предусматривающему участие электронов обоих
атомов, но и по иному механизму, называемому донорно-акцепторным: один атом
(донор) предоставляет для образования связи неподеленную пару электронов, а
другой (акцептор) – вакантную квантовую ячейку.
Вне
сомнений, молекулы, полученные по разным механизмам, неразличимы; связи
совершенно равноценны. Поэтому правильнее не выделять донорно-акцепторное
взаимодействие в особый вид связи, а считать его лишь особым механизмом
образования ковалентной связи.
Связь,
образованная перекрыванием электронных облаков по оси, проходящей через ядра
атомов, называется s-связью.
Если
в связи участвует s-орбиталь, то может образоваться только s-связь. Отсюда она
и получила свое название, т. к. греческая буква s является синонимом латинской
s.
При
участии в образовании связи р-орбитали и
d-орбитали перекрывание по s-типу
осуществляется в направлении наибольшей плотности электронных облаков, которое
и является наиболее энергетически выгодным. Поэтому при образовании связи такой
способ всегда реализуется в первую очередь. Следовательно, если связь
одинарная, то это обязательно s-связь, если кратная, то одна из связей
непременно s-связь.

Примеры s-связей
Однако
из геометрических соображений понятно, что между двумя атомами может быть
только одна s-связь. В кратных связях вторая и третья связи должны быть
образованы другим геометрическим способом перекрывания электронных облаков.
Вследствие
существования химической связи и перекрытия электронных облаков атомов,
составляющих молекулу между ними возможно возникновение запутанных состояний.
Для простейшей молекулы водорода существование запутанных состояний очевидно.
Для более сложных соединений необходимы дополнительные исследования с
привлечением аппарата квантовой химии.
Но как может возникать запутывание в
между электронными облаками молекул разных клеток и ( что для данного
рассмотрения важно ) различных организмов?
Рекомбинация
генетическая - реорганизация генетического материала,
обусловленная обменом отдельными сегментами (участками) двойных спиралей ДНК.
Различают
два основных типа генетической
рекомбинации: 1) «законную» (общую, или гомологичную), при которой
происходит обмен гомологичными (одинаковыми) участками молекул ДНК; 2) «незаконную»
(негомологичную), в основе которой лежит обмен негомологичными участками ДНК.
Законная
генетическая рекомбинация наблюдается, например,
между двумя копиями какой либо хромосомы. У эукариот (все организмы, за
исключением бактерий и синезеленых водорослей) наиболее типичен обмен участками
гомологичных хромосом в мейозе (деление клеток, в результате которого
происходит уменьшение числа хромосом в дочерних клетках-основная стадия
образования половых клеток). Этот обмен может происходить между плотно
конъюгированными хромосомами на ранних стадиях развития яйца или сперматозоида.
Реже законная генетическая рекомбинация осуществляется при обычном делении
клеток (с сохранением числа хромосом)-митозе.
В
основе молекулярного механизма законной генетической
рекомбинации лежит принцип «разрыв-воссоединение» двух гомологичных
молекул ДНК. Этот процесс (его наз. кроссинговер) включает несколько промежуточных
этапов: 1) узнавание участков; 2) разрыв и реципрокное (крест-накрест) воссоединение
молекул: замена одних цепей гомологичными; 3) устранение ошибок, возникающих в
результате неправильного спаривания участков. Точка обмена может возникать на
любом участке гомологичных нуклеотидных последовательностей хромосом,
вовлекаемых в обмен. При этом в точке обмена обычно не происходит изменения
нуклеотидных последовательностей. Точность разрыва и воссоединения чрезвычайно
велика: ни один нуклеотид не утрачивается, не добавляется и не превращается в какой-то
другой.
Основой всех предложенных схем генетической рекомбинации послужила так называемая модель Холлидея,
согласно которой генетическая рекомбинация начинается с
разрыва только одной из двух цепей спирали ДНК. Вслед за разрывом один конец
цепи вытесняется другим концом, который
наращивается ДНК-полимеразой. Вытесненный конец разорванной цепи спаривается со
второй молекулой ДНК (образуется так называемый гетеродуплекс), в свою очередь
вытесняя там участок одной из ее цепей. В конце концов одиночные гомологичные
цепи обмениваются реципрокно. После этого первоначального этапа спаривания две
гомологичные спирали ДНК удерживаются вместе благодаря перекрестному обмену
цепями по одной от каждой спирали. Точка перекрестка далее может мигрировать, в
результате чего дополнительно образуются или растут гетеродуплексные участки на
обеих молекулах ДНК.
Структура с перекрещенными цепями может
существовать в различных стереоизомерных формах, возникающих в результате
вращения составляющих ее элементов относительно друг друга. Изомеризация, которая
как и другие стадии генетической рекомбинации контролируется генетически, изменяет положение двух пар цепей: две ранее
перекрещивавшиеся цепи становятся неперекрещивающимися и наоборот.
Для
того чтобы вновь восстановились две отдельные спирали ДНК и тем самым прекратился
процесс спаривания, в каждой из двух перекрещенных цепей должен произойти
разрыв. Если он происходит до того, как прошла изомеризация, то две исходные
спирали ДНК отделяются друг от друга так, что у каждой из них генетически
перестроенной оказывается только одна цепь. Если же разрыв двух перекрещенных
цепей происходит после изомеризации, то обе молекулы ДНК претерпевают полную
реорганизацию: часть каждой исходной спирали оказывается присоединенной
(ступенчатым соединением) к части другой спирали.
Законная
генетическая рекомбинация приводит к возникновению новых комбинаций
специфических аллелей (различной формы одного и того же гена, обусловливающие
различные варианты развития одного и того же признака-группы.

Текущая версия (не проверялась)

Иллюстрация
кроссинговера, Томас Хант Морган (1916)
Кроссинго́вер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе
[33]. Помимо мейотического описан также митотический
кроссинговер.
Кроссинговер происходит в профазе I
мейоза во время конъюгации гомологичных хромосом. В это время части двух
хромосом могут перекрещиваться и обмениваться своими участками. В результате
возникают качественно новые хромосомы, содержащие участки (гены) как материнских,
так и отцовских хромосом. Особи,
которые получаются из таких гамет с новым сочетанием аллелей, получили название
кроссинговерных или рекомбинантных.

Конъюгация
Конъюгация (от
лат. conjugatio - соединение). Конъюгация хромосом -
попарное временное сближение гомологичных хромосом, во
время которого между ними может произойти обмен гомологичными участками. После конъюгации хромосомы
расходятся.
В силу механизма гомологичной генетической
рекомбинации, частным случаем которой является механизм кроссинговера, при
перекресте и дальнейшем расхождении хромосом генетический материал различных хромосом
может быть связан посредством запутанных состояний.
Но может ли существовать такая связь
между хромосомами различных клеток ?
Митоз
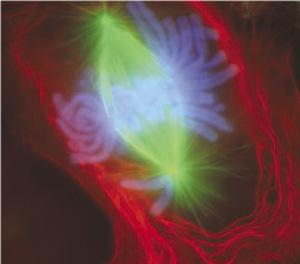
Ранняя анафаза митоза в клетке почки тритона (световой микроскоп,
иммунофлюоресценция). Микротрубочки зелёные, хромосомы голубые.
Митоз (реже: кариокинез или непрямое
деление) — деление ядра эукариотической клетки с сохранением числа хромосом [33].
Как
видно хромосомы расходятся по разным клеткам, что обеспечивает нелокальную
связь между последними, посредством запутанных состояний.
Здесь,
однако, следует сделать отдельное замечание.
Митотический (соматический) кроссинговер. Мейотический
кроссинговер имеет место практически всегда. Что же касается гомологичной
генетической рекомбинации в ходе деления соматических клеток, то она происходит
реже. В рамках рассматриваемой гипотезы важно,
чтобы передача информации осуществлялась в период эмбриогенеза и касалась
центральной нервной системы, так как
ее клетки существуют на протяжении всей жизни организма и не подвержены делению
по завершении указанного процесса.
Кроссинговер,
как уже упоминалось, может происходить не только во время мейоза, но и митоза –
в соматических клетках, тогда его называют митотическим. В этом случае он
приводит к формированию мозаичных признаков.
Соматический
кроссинговер может быть обнаружен, если он осуществляется на стадии четырех
хроматид. Гомологичные хромосомы в интерфазе конъюгируют и входят в
митотическое деление спаренными.
Частота митотического кроссинговера
значительно реже мейотического. Тем не менее его также можно использовать для
генетического картирования.
Для возникновения физиологически значимого
количества запутанных состояний совсем не обязательно, чтобы в процессе каждого
митотического деления имел место кроссинговер. Учитывая, что количество клеток
в центральной нервной системе измеряется десятизначным числом, достаточно,
чтобы лишь определенный процент клеток в процессе эмбриогенеза образовывался с
включением механизма гомологичной генетической рекомбинации. В этом случае
кора, подкорковая структура и некоторые другие образования мозга смогут
образовать каркас, на котором согласуюясь с голографическим принципом К.
Прибрама будет существовать сознание.
Но это – только в рамках одного
организма.
Мейоз (или редукционное
деление клетки) — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза [33].
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от
диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной)
происходит в результате полового процесса.
Таким образом, следует
констатировать возможность
существования запутанных состояний между генетическим материалом родительских и
дочерних особей.
Такого рода
запутанные состояния могут существовать на всех уровнях биологического объекта,
включая центральную нервную систему.

Но только ли непосредственно
между родителями и потомством будет возникать квантовая сцепленность? Отнюдь
нет. Хотя социальные нормы запрещают имбридинг все равно все мы, с точки зрения
молекулярной генетики в той или иной степени – дальние родственники.
Ученый Ф. Вайденрайх сделал доклад в Стокгольме в 1938 г. на II Международном
конгрессе антропологических и этнографических наук. Позже он дополнительно
аргументировал свою гипотезу в целом ряде фундаментальных трудов. Она названа
была им полицентрической, так как постулировала независимое возникновение
современного человека в нескольких центрах.
Через
некоторое время после публикации полицентрических работ Ф. Вайденрайха Я.Я. Рогинским была возрождена так называемая
моноцентрическая гипотеза в форме гипотезы широкого моноцентризма.
Итак, нельзя исключать существование
квантовой сцепленности (запутанности) электронных облаков молекул ДНК большого
количества субъектов.
Вообще
говоря, данное предположение наталкивалось на серьезные препятствия, и долгое
время могло считаться неверным по следующей причине.
Дело
в том, что в квантовой физике детально изучается явление декогеренции – процесса исчезновения квантовомеханических свойств
системы ( отличающейся от свойств наблюдаемых в макромире ) при воздействии
различного рода факторов, главным образом электромагнитных полей.
Однако детальное рассмотрение данного
вопроса в ракурсе интерпретации квантовой механики, основанной на гипотезе «относительных состояний», выдвинутой Хью Эвереттом в
1957 году, привело
к обнадеживающим выводам. [30] В результате
процесса декогеренции не происходит исчезновение запутывания, а имеет место
усложнение последнего. При воздействии фотонов (а именно они, как упомянуто
выше, являются основными факторами декогеренции) наблюдателю становится
проблематично фиксировать все составляющие системы. При исключении из
рассмотрения квантов электромагнитного поля, с околосветовыми скоростями
уходящими из области наблюдения, теряется информация о фазах комплексных
коэффициентов входящих в описание волновых функций.
«Узкие» места
гипотезы
Как
и каждая «объемная» теория, изложенная гипотеза имеет ряд положений, доказать
которые весьма проблематично и, которые, вследствие этого, являются
потенциальной мишенью для справедливой критики оппонентов. Приведу, на мой
взгляд, наиболее «проблемные» вопросы.
1. Проблема декогеренции. Конечно же,
это один из важнейших аспектов теории. Несмотря на рассмотрение данного
вопроса, в ракурсе гипотезы Хью Эверетта, тот факт, что запутанные состояния
продолжают существовать длительное время, нуждается в экспериментальной
проверке. Экспериментальные работы, в рамках исследований феномена
декогеренции, в настоящее время проводятся в различных научных центрах мира.
2. Запутанные состояния электронных
облаков. Как было отмечено, вопрос о
возможности возникновении запутанных состояний между электронными облаками
атомов водорода при разрыве химической связи молекулы, не вызывает сомнений.
Что же касается более сложных молекул, необходимы дополнительные исследования с
привлечением аппарата квантовой химии.
3. Генетическая рекомбинация. Обмен
гомологичными участками хромосом при кроссинговере ( гомологичная генетическая
рекомбинация ) осуществляется посредством механизма Холлидея с образованием
соответствующих структур. Действительно ли они обеспечивают такой обмен между
нитями нуклеиновых кислот, что при делении клетки обе копии генетического
материала связанны квантовой нелокальностью – является вопросом.
Вообще
говоря, автор настоящей работы не видит принципиальных ограничений на
возможность существования описанного механизма ( концептуальным, правда
является вопрос декогеренции ). Реализовала ли Природа подобную возможность ? Ответ на этот вопрос сможет дать только
эксперимент.
Следует
отметить, что приведенная в данном исследовании гипотеза не совсем нова и
уникальна. Возникновение запутанных состояний между генетическим материалом
рассматривалось одним из российских исследователей (кандидат биологических наук,
специалист в области молекулярной генетики Петр Петрович Гаряев), но только в
рамках одного организма. Причем автору настоящей статьи не удалось понять, каким
именно образом гипотеза П.П. Гаряева определяет возникновение квантовой
нелокальности – там речь не идет о перекрытии электронных облаков и механизме
кроссинговера. А без механизма гомологичной генетической рекомбинации едва ли
удастся решить проблему – водородные связи, посредством которых «сшита» двойная
спираль ДНК, вряд ли могут нести физиологически значимую информацию. Ученым из
Великобритании также специалистом в области молекулярной генетики Рупертом
Шелдрейком выдвигалось предположение о существовании связи между генетическим
материалом различных особей посредством некоего физического поля, но не
определялось какого именно ( возможно потому что Р. Шелдрейк не является
физиком ). А другим российским ученым – физиком Василием Леонидовичем Янчилиным
постулировалось наличие запутанных состояний между биологическими объектами, но
никак не конкретизировалось, посредством какого механизма происходит их
передача потомкам.
Нельзя
не отметить что подобное изыскание может пролить свет на так называемую
психофизиологическую проблему – вопрос о соотношении физиологических структур (
подразумевается мозг ) и психической сущности.
Разумеется,
изложенное в данном исследовании должно подвергнуться тщательному критическому
анализу и, по возможности, должно быть подтверждено экспериментально. Но
существование направления этих изысканий, на взгляд автора, вполне правомерно и
должно заполнить свою нишу в сфере научной деятельности, даже с учетом
возможного дальнейшего опровержения. Как говорится «отрицательный результат –
тоже результат».
Литература
- Артюхов В.Я., Майер Г.В. Квантово
- химическая теория переноса энергии электронного возбуждения в
молекулярных системах// Журнал физической химии. - 2001. -Т.75. - №6. -
С.1143-1150.
- Амусья М.Я., Иванов В. К.
Межоболочечное взаимодействие в атомах // Успехи физических наук. – 1987.
- № 2.- С. 185-230.
- Александров А.А. Психогенетика.-
СПб.: Питер, 2004. – 192 с.
- Астраментова Л. Введение в
психогенетику. – М.:МПСИ, 2004.-472 с.
- Баргатин И.В., Гришанин Б.А.,
Задков В.Н. Запутанные квантовые состояния атомных систем // Успехи
физических наук. – 2001. - № 6. – С. 625-647.
- Белинский А.В. Квантовая
нелокальность и отсутствие априорных значений измеряемых величин в
экспериментах с фотонами // Успехи физических наук. – 2003.
- Бом Д. Квантовая теория. М.,1965.
- Буренин А.В. Симметрия квантовой
внутримолекулярной динамики // Успехи физических наук. - 2002. - № 7. – С. 813-836.
- Бучаченко А.Л. Химическая
поляризация электронов и ядер. - М.. Наука, 1974, 245 с.
- Вальков В.В. Применение теории
групп в квантовой химии. – Красноярск: КГУ, 1994, 47 с.
- Введение в квантовую химию./
Накагура С., Накадзима Т., Енедзава. - М.: Мир, 1982. - 264 с.
- Гриб А.А. Неравенства Белла и
экспериментальная проверка квантовых корреляций на макроскопических
расстояниях // Успехи физических наук. – 1984. - № 4. – С. 619-634.
- Cтанислав Гроф. «За пределами
мозга: рождение, смерть и трансценденция в психотерапии». М.,
1994.
- Гэйто Дж. Молекулярная
психобиология. - М.: Мир, 1969. - 276с.
- Дяткина М.Е., Основы теории
молекулярных орбиталей. - М.Наука, 1975.
-189с.
- Дьюар М., Теория молекулярных
орбиталей в органической химии. - М.: Мир. 1972. - 590 c.
- Попл А. Джон. Квантово-химические
модели. Нобелевская лекция. Стокгольм, 8 декабря 1998 г.
- Зельдович Я.Б., Бучаченко А.Л.,
Франкевич Е.Л. Магнитно-спиновые эффекты в химии и молекулярной физике //
Успехи физических наук. -1988.-№ 1.- С. 3-45.
- Квантовая биохимия./ Пюльман Б.,
Пюльман А. - М.: Мир, 1965. - 654 с.
- Квантовая механика молекул. /
Мак-Вини Р., Сатклиф Б. - М.: Мир, 1972. -
380с.
- Квантовая механика молекул и
квантовая химия. Учебн.пособие./ Степанов Н.Ф., Пупышев В.И. - М.: Изд-во
Моск. ун-та, 1991.- 384 с.
- Квантовая органическая химия./ К.Хигаси,
Х.Баба, А.Рембаум. - М.: Мир.
- Квантовая химия. / Л.А. Грибов, С.П.
Муштаков. – М.: Гардарики. – 1999. – 390 с.
- Квантовая химия органических
соединений. Механизмы реакций. / Минкин В.И., Симкин Б.Я., Миняев Р.М. - М.:
Химия. 1986. - 248 с.
- Клышко Д.Н. Парадокс
Эйнштейна-Подольского-Розена для наблюдаемых «энергия-время» // Успехи
физических наук. – 1989.-№1.- С. 327-341.
- Краснов К.С. Молекулы и химическая
связь: Учеб. пособие. 2-е изд. - М.: Высш. шк., 1984. - 275 с.
- Ладин Я. Квантовая биохимия для
химиков и биологов. - Москва: «Мир», 1975. - 252 с.
- Межконтинентальные квантовые связи
между запутанными электронами в ионных ловушках термолюминисцентных
кристаллов / Роберт Дебранд, Дэниел Л. Ван Гент (quant-pb/06 11109)
- Менский М.Б. Квантовое измерение:
декогеренция и сознание // Успехи физических наук.-2001. - № 4.-C. 459-462.
- Менский М.Б. Концепция сознания в
контексте квантовой механики // Успехи физических наук.-2005. - № 4.-C. 413-435.
- Модель отталкивания электронных
пар валентной оболочки и строение молекул: Пер. с. англ./ Гиллеспи Роналд,
Харгиттаи Иштван – М.: Мир, 1992. – 296 с.
- Молекулярная генетика./ Стент Г.,
Кэлиндар Р. - М.: Изд. Мир, 1981. - 648 с.
- Нокаряков. Хромосомные и
молекулярные основы наследственности. - М., 1977 - 88 с.
- Нуклеиновые кислоты мозга./ Лукаш
А.И., Короленко П.И. – Ростов н/Д: из-во Рост. ун-та, 1973. – 20 с.
- Основы квантовой химии. /
Заградник Р., Полак Р. - М.: Мир, 1979. - 504 с.
- Основы психогенетики. / Малых
С.Б., Егорова М.С., Мешкова Т.А.-М.:Эпидавр.-1998.-744 с.
- Петров Р.В. Иммуногенетика и
искусственные антигены. - М.: Медицина, 1983. - 256 с.
- Прибрам К., "Языки мозга:
экспериментальные парадоксы и принципы нейропсихологии", М., 1975.
- Психогенетика. / И.В. Равич-Щербо,
Т.М. Марютина, Е.Л. Григоренко; Под ред. И.В. Равич-Щербо. -М.: Аспект
Пресс, 2000.-447 c.
- Рунов Н.Н. Строение атомов и
молекул. - М.: Просвещение, 1987. - 144 с.
- Слэтер Дж., Электронная структура
молекул, пер. с англ., - М., 1965
- Счастнев П.В. Теория электронных
оболочек молекул. – Новосибирск.: НГУ, 1973. -143с.
- Сент-Дьерди А. Введение в
субмолекулярную биологию. - М.: «Наука», 1964. - 139 с.
- Теория возмущенных молекулярных
орбиталей в органической химии. / Дьюар М., Догерти Р. - М.: Мир, 1977. -
695 с.
- Теория групп и ее применение в
квантовой механике молекул./ Болотин А.Б., Степанов Н.Ф. - М.: Изд-во
Моск. ун-та, 1973. - 227 с.
- Теория электронного строения
молекул./ Мулдахметов З.М., Минаев Б.Ф., Безносюк С.А. - Алма-Ата: Наука.
1988. – 216 c.
- Уотсон Дж. Молекулярная биология
гена. - М.: Мир, 1967. - 464 с.
- Физика квантовой информатики./
Боумейстер Д., Экерт А., Цайлингер А. - М.: "Постмаркет", 2002.
- 376 с.
- Фларри Р. Квантовая химия. - М.:
Мир, 1985. - 472 с.
- Франк-Каменецкий М.Д., Аншелевич
В.В., Лукашин А.В. Полиэлектролитная модель ДНК // Успехи физических
наук.-1987.-№4.-С.595-618.
- Франк-Каменецкий М.Д. Топология и
структурные переходы в ДНК // Успехи физических наук.-1986.-№4.-С.729-732.
- Хабердитцл В. Строение материи и
химическая связь: Пер. с нем. – М.: Мир, 1974. – 296 с.
- Хачатрян Г.С. Биохимия нуклеиновых
кислот и высшие функции головного мозга. - Ереван: Айтастан, 1981. - 332
с.
- Холево. Введение в квантовую
теорию информации. - М.: МЦНМО, - 2002. - 128 с.
- Элементарное введение в квантовую
биохимию./ Голованов И.Б., Пискунов А.К., Сергеев Н.М. – М.: Наука,
1969.-235с.
- Юнг К.Г. Синхрония. Москва – Киев:
Релф-бук – Ваклер, 2003. - 320 с.
- Penfield W. The mystery of the mind. — 2-d ed. — Princeton; New
Jersey: Princeton University
Press, 1978. — 123 p.
- Popper K.R., Eccles J.C. (1977) The Self and Its Brain.
Springer International, Berlin,
Heidelberg,
London: New York.

